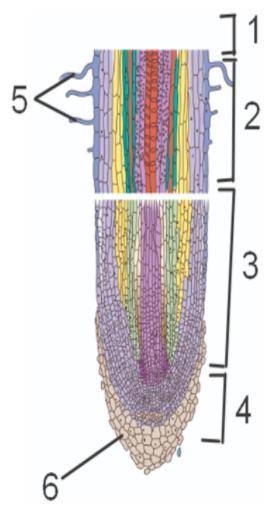
1. Напишите названия зон корня, указанных на рисунке цифрами 1–4. 1. 2. 3. 4.
2. Каково значение корневого чехлика? Запишите цифру, которой он обозначен на рисунке?
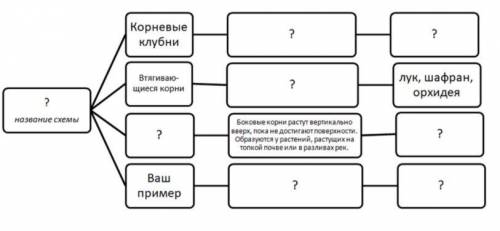
Задание 2. Рассмотрите приведённую ниже схему. Дайте название схеме, дополните её необходимыми данными, примерами растений – запишите конкретные названия и выполните схематические рисунки или прикрепите готовые иллюстрации. (файл 2)
Дорогой ученик,
Признаки, которые характерны для головоногих моллюсков, именно три, это:
1) Ротовое отверстие окружают две толстые роговые челюсти (пункт Б) - это один из признаков, отличающих головоногих моллюсков от других групп моллюсков. Челюсти помогают им захватывать и измельчать пищу перед тем, как она попадает в желудок.
2) Сердце двухкамерное (пункт Д) - головоногие моллюски имеют сердце, состоящее из двух камер. Это отличает их от более примитивных моллюсков, у которых сердце может иметь одну камеру.
3) В жизненном цикле имеется стадия личинки (пункт Е) - головоногие моллюски проходят через стадию личинки, которая отличается от взрослой особи по строению и образу жизни. Эта стадия позволяет им приспосабливаться к определенным условиям водной среды.
Давай теперь рассмотрим остальные варианты и почему они неверные:
- Почти замкнутая кровеносная система (пункт А) - это признак общий для всех моллюсков в целом и не помогает нам отличить головоногих моллюсков от других моллюсков.
- Имеют органы воздушного питания – легкие (пункт В) - это признак характерный для некоторых других классов моллюсков, например, у некоторых моллюсков есть легкие. Однако у головоногих моллюсков легких нет, они дышат при помощи жабер.
- Нервные узлы очень велики и слиты в крупное образование – головной мозг (пункт Г) - это признак позвоночных (хордовых) животных, а не моллюсков.
Таким образом, признаки, характерные для головоногих моллюсков, включают в себя ротовое отверстие окружают две толстые роговые челюсти, двухкамерное сердце и наличие стадии личинки в жизненном цикле.
Надеюсь, ответ был понятен и полезен. Если у тебя еще есть какие-то вопросы, не стесняйся задавать!"
Кальмар - это морское животное, относящееся к классу головоногих моллюсков. Для того чтобы определить признаки, которые НЕ присущи кальмару, мы должны посмотреть на перечисленные характеристики и исключить те, которые справедливы для кальмара.
1. Тип питания - хищничество: Кальмар является хищником и питается рыбой, поэтому этот признак присутствует.
2. Живет в морской воде: Кальмары обитают в морской воде, поэтому этот признак также присутствует.
3. Мягкое нечленистое тело, наличие мантии: У кальмара тело мягкое и нечленистое, а мантия является его основной частью. Поэтому этот признак также присутствует.
4. Наличие двустворчатой раковины: Кальмары не имеют раковину, поэтому этот признак НЕ присущ кальмару.
5. Глаза расположены между основанием рук и щупальцев: У кальмара глаза расположены на передней части тела, поэтому этот признак НЕ присущ кальмару.
6. Наружная раковина редуцирована: Кальмары не имеют наружной раковины, поэтому этот признак присутствует.
7. Раковина спирально закручена, имеет вид башенки: Кальмары не имеют раковины, поэтому этот признак НЕ присущ кальмару.
8. Тип питания - фильтратор: Кальмары являются хищниками и не фильтруют пищу, поэтому этот признак НЕ присущ кальмару.
9. Реактивное передвижение: выталкивание воды из мантийной полости: Кальмары передвигаются, выталкивая воду из мантийной полости, поэтому этот признак присутствует.
10. Орган дыхания - "легкое": У кальмара нет легких, его органы дыхания - жабры. Поэтому этот признак НЕ присущ кальмару.
11. Органы дыхания - жабры: У кальмара жабры являются органами дыхания, поэтому этот признак присутствует.
12. Практически замкнутая кровеносная система: У кальмаров кровеносная система замкнутая, поэтому этот признак присутствует.
13. Глаза редуцированы: У кальмара глаза находятся на передней части тела и не являются редуцированными, поэтому этот признак присутствует.
14. Растительноядный: Кальмары хищник и не является растительноядным, поэтому этот признак НЕ присущ кальмару.
15. По бокам головы расположены крупные глаза: У кальмара глаза находятся на передней части тела и не расположены по бокам головы, поэтому этот признак НЕ присущ кальмару.
Таким образом, признаки, которые НЕ присущи кальмару, это:
4. Наличие двустворчатой раковины.
5. Глаза расположены между основанием рук и щупальцев.
7. Раковина спирально закручена, имеет вид башенки.
8. Тип питания - фильтратор.
10. Орган дыхания - "легкое".
14. Растительноядный.
15. По бокам головы расположены крупные глаза.